

When I was a callow teenager in the 1970s, studying Biological Sciences at the University of Edinburgh, our lecturers had an aphorism: “The law recognises two sexes: science recognises seven”. And we’ve found a few more since then
Yet, anybody who has been on the net for any length of time has seen people claiming that “science says” that there are only two sexes, determined by whether you have two X chromosomes, or an X and a Y. Even leaving aside for the moment that this has only ever been applicable to marsupial and placental mammals and a few outliers such as the Gingko biloba tree, it’s waaaayyyy more complicated than that. Like molecular valencies, the binary XX/XY split is what we call “Lies to children” – a “good enough” approximation we teach to small children because it gets them into the right general area of knowledge, but which is grossly over-simplified.
The making of a man (a cis man, anyway)
In mammals all embryos start out with a basic blueprint for a female body plan, and with neutral rudimentary gonads. They then develop male characteristics if certain conditions are met, and if not, they keep the original body plan and develop ovaries. In therian mammals (marsupials and placentals) these conditions are:
- Possession of an SRY (Sex-determining Region Y) gene. This is usually but not always sited on a Y chromosome.
- The SRY gene functions normally and triggers the development of testicles instead of ovaries.
- The testicles function normally and produce a quantity of androgens sufficient to trigger the development of male characteristics, assuming that the correct genes are present on the X chromosome (see below).
- Possession of the correct gene variants on the X chromosome to enable the tissues of the embryo’s body to be normally responsive to androgens. If these are absent or damaged, the embryo will have Androgen Insensitivity Syndrome (AIS), which comes in varying degrees of severity.
- In Mild AIS the person appears to be a fairly normal male, but is sterile.
- In Partial AIS the child will be born with a mix of male and female characteristics, or may be born apparently entirely female, but then develop a penis and scrotum at puberty in response to a sudden hormon surge. There’s a community in the Dominican Republic where so many children are born with this syndrome that they have a special local name – they’re called Guevedoces or “penis-at-twelves”.
- In Complete AIS, the child will be born externally female in all ways, but will lack a uterus and have internal, undescended testicles instead of ovaries. Often such people live their whole lives believing themselves to be simply a sterile woman. The reason it used to be common to peform sex tests on female athletes was because the Soviet Union had discovered that such women had a degree of male muscle development, and was collecting them and using them to cheat at international sports.
- Also, possibly, not be exposed to very large amounts of female hormones during development.
If all these conditions are met, the person will be at least potentially a genetically and anatomically normal male, but there are many other ways a person can end up as an interesex.
Freemartins and other sexual intermediates
The following are some of those more-than-seven sexes which can occur in therian mammals – and I’ve probably forgotten a few. Most result in what are called “intersexes” – individuals whose physical sexual characteristics are either intermediate between male and female, or an assortment of male and female characteristics.
Genetically and developmentally normal female with XX chromosomes and ovaries.
Genetically and developmentally normal male with XY chromosomes and testicles.
Persons with Androgen Insensitivity Syndrome (see above), with XY chromosomes but no penis or scrotum, with a vagina, clitoris and breasts but with internal testicles instead of ovaries, and no womb. Some grow a penis and scrotum at puberty, some never do.
XY Gonadal Dysgenesis or Swyer Syndrome – the person has XY chromosomes but is fully female, externally and internally, except that in the most extreme form they have neither testicles nor ovaries: just a smear of unformed gonadal tissue known as a streak gonads which is usually removed as it is prone to becoming cancerous. Usually due to loss of or damage to the SRY gene. Note that some XY women have been found to be fertile and able to get pregnant, and in mice it is common for XY females to be fertile, although they should on average have a 2:1 ratio of female to male babies, rather than 1:1.
There is also a condition called XX Gonadal Dysgenesis, where the person has normal female XX chromosomes and female anatomy but has streak gonads instead of ovaries, much like a person with Swyer Syndrome. Occasionally, gonadal dysgenesis can leave a mostly-female person with ovotestes – gonads which combine the features and function of both ovaries and testicles – although this is so rare in humans we don’t really seem to know why it happens. In moles, for some reason, it’s the norm.
XX Male or de la Chapelle Syndrome – persons who are XX but are in all or most respects male, with testicles, but sterile. Usually this is due to a genetic crossover between X and Y which resulted in the SRY gene ending up on an X chromsome, but there are also much rarer SRY-negative XX males, occurring due to a mutation of a gene already present on an X chromosome.

MBLWHOI Library Rare Book Collection
Freemartins – persons who are XX and would have been female but have intermediate external genitalia and no uterus, because they were exposed to an excess of male hormones in the womb. This is seen very rarely in humans and occasionally in sheep, goats and mice, but is very common in cattle. The usual cause is that male and female fraternal twin embryos implanted side by side and their placentas became linked, so that androgens from the male twin’s testes leaked through into the female embryo. For some reason the placentas of cattle are especially prone to this. “Freemartin” is such a nice word that I personally think it should be borrowed by human intersexes in general.
Hypospadias (displacement of the urethral opening away from the tip of the penis), cryptorchidism (undescended testicles) and other genital abnormalities in persons who are XY and are otherwise genetically normal males. These abnormalities are becoming increasingly common and it is theorised that this is due either to pollution with “endocrine disrupting” industrial chemicals, or to an increasingly fatty diet leading to a rise in oestrogens in the womb. However, the evidence for this is inconclusive. There are also people who are genetically and internally female, but are born with either an over-large clitoris that looks like a penis, or with an actual fully-developed penis (that is, with no vaginal opening and with the urethra and vagina venting through the penis: this is the norm in spotted hyaenas but also sometimes happens in humans).
Chromosomal disorders of the sex chromosomes, in which the individual ends up with more or fewer sex chromosomes than the usual two (there are other chromosome disorders such as Down’s Syndrome which occur when the person ends up with more or less than two of one of the autosomal, non-sexual chromosomes). Every cell (except red blood cells and gametes) in a chromosomally normal therian mammal has two of each sort of chromosome, one from each parent. During meiosis, the production of gametes, the pairs of chromosomes line up together and double up so there are two pairs of them instead of two individual chromosomes, and then exchange some of their genes, so each chromosome ends up with some genes which came originally from the person’s mother and some from their father. Then they split up to form four separate chromosomes and one of each goes to each gamete, so each gamete ends up with one copy of every type of autosomal chromosome, and one sex chromosome – in theory.
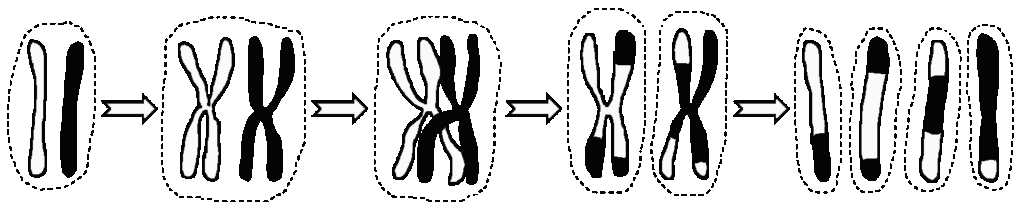
However, it’s fairly common for this process to go wrong, so that some gametes end up with none of a particular chromosome, and some end up with two. When this happens with the sex chromosomes, you get intersexes. Possibly the commonest is Klinefelter’s Syndrome, XXY males, who are slightly feminised (wide hips, extra breast tissue etc) and have reduced fertility, and for some reason unusually mobile elbows. The extra X may come from either the mother’s or the father’s gametes. You can also get XXXY males and other multiples. XYY Syndrome males tend to be tall and strong with an increased risk of acne and of learning disabilities. Women with Turner’s Syndrome, who have a single X chromosome and no Y, tend to be short and sub-fertile with an increased risk of heart disease and may have gonadal dysgenesis (failure of the ovaries to develop properly). There’s no male equivalent: there are genes on the X chromosome which we need to produce a viable mammal embryo and which have been lost from the Y, so a zygote with a Y chromosome and no X won’t get past being a small ball of cells.

Then you have mixed sex chimaeras. Chimaeras form when fraternal twin embryos fuse in the womb, generating an embryo which is a mix of two different genetic lines. If the initial embryos were of different sexes the final embryo will be an intersex. Some will be a hodgepodge of different parts, while others will be divided neatly down the middle or across the waist. This is most easily seen in flowers, or in birds of species where the sexes are different colours. But it happens in mammals too – in fact one of my university textbooks had a photograph of an unfortunate black South African who was male down one side and female down the other. It even affected their face, which had a larger jaw and heavier brow on the male side.

It would also be possible to get a mixed-sex individual who was a mosaic, although I don’t know if this has ever been observed. Mosaics occur when one or more cells mutate, in a growing foetus which has a single genetic origin. If it happens late in development you just get a small patch of mutant cells, such as a white patch in the hair, but if it happens at the two-cell stage the resultant individual will be a 50:50 mix of the two cell lines. It would be possible for one of the cells in an XY male to lose the Y and generate X0 female cells, mixed in with the male ones.
Sex versus gender
Biological sex and gender are connected, but not synonymous. Gender has to do with how you feel and how you behave, but people who are transgender generally feel strongly that they are not only behaviourally different from their apparent physical sex at birth, but that their bodies are wrong and need to be alterred to fit the physical type which is most commonly identified with the gender they feel they are.
I have trouble visualising what it must be like to be transgender, because I have trouble imagining what it would be like to feel like a sex or gender in the first place. I feel like a me. If I woke up tomorrow and I was a bloke, then aside from the initial “How the fuck did that happen?” I doubt if it would take me more than a few days to adjust.
There are also people like Barry Humphries and Grayson Perry and my late friend Jonathan Waite, who are cis gender (that is, the gender that most often matches their biological sex) most of the time, but have an alter ego who is a different gender, who appears to be not just a performance but an outgrowth of their personality, but who is only active part-time.
I have various half-formed theories about what might be going on here, spiritual, sociological and medical. The spiritual one, which is meaningful to me as a believer in reincarnation, is “Maybe such people have been the other sex for many lives and haven’t quite made the transition”. But we can set that aside as currently untestable, and also not necessarily making a difference in re. the other ideas.
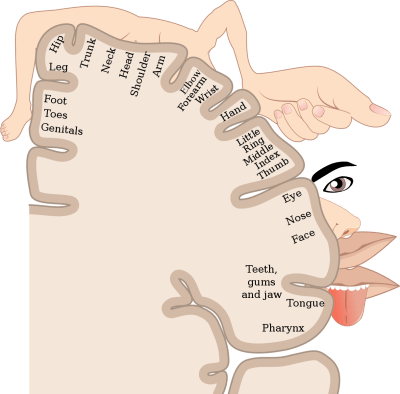
The medical one is that, for whatever reason, the person’s cortical homunculus – the part of the brain that maps onto various body parts – doesn’t match their sexual characteristics. That would cause the sexual parts that they do have to feel foreign, in the same way that some kinds of brain damage can make somebody feel that their arm belongs to somebody else.
It’s not as simple as that, though, because then you have to ask why the cortical homunculus wouldn’t match the body. It seems to me that transgender people often look a lot more like the sex they identify as, even before they begin to consciously model themselves as that sex, which suggests that at a hormonal and developmental level they were always the gender they identify as, or at least heading that way, and that’s why their cortical homunculus developed the way it did (if it did). I think for example of an M2F trans woman I knew in London in the 1980s who, OK, made a very slightly mannish woman, as she had large hands and Adam’s apple, but she must have made an unbelievably girly boy before she transitioned, and I never had the slightest doubt that she was a she. I’ve met born, cis women who were far more masculine.
For this reason, I tend to see fully transgender people as another sort of intersex – but as intersexes nearly all of whom are far more like the sex they feel they are than the one they were identified as at birth, so that it makes perfect sense for them to be alterred so their body matches their inner map. Even if it were possible to change their brain map to match their body instead, that wouldn’t be as good a solution because most appear to already have physical/hormonal elements of the sex they identify with, even before they transition.
[Update as at 2023: the latest research suggests that transgender people do indeed have the neurological wiring of the sex they feel themselves to be, probably due to hormone fluctuations in the womb, so their brain’s map of their body doesn’t match or feel comfortable with the sexual parts they were born with. But at present we can’t prove that all transgender people show this feature, because at present it’s only identifiable by post-mortem dissection.]
At the same time, I do wonder whether as many people would be transgender if society stopped telling people that this or that behaviour was typically masculine or feminine, and just let them get on with liking whatever they like. Also I wonder about those Native American societies which accepted that you were whatever sex/gender you were inside your head, so that it was perfectly acceptable to be e.g. a woman with a dick or a man with a bust. If you were a transgender woman in such a society, did you still obsess over wanting to get rid of your male-type genitalia, or did you feel comfortable with being a woman who had a dick, because that was an acceptable way of being a woman in your society? If the answer is yes, they still hated having male genitalia, that would support the cortical homunculus hypothesis, and if not, not.
Sexual determination in other organisms
The above applies to nearly all placental and marsupial mammals, whose system of XY chromosomes and an SRY gene is very stable and seems to have been in place for about 150 million years. However, there are a whole lot more different ways of doing sex and gender in other organisms: and even in therian mammals, some rodents have lost the Y-chromosome altogether, while others sometimes have fertile XY females alongside “normal” XY males and XX females.

Monotremes (one species of duck-billed platypus and four of echidna) are also mammals, but sexually they’re a whole other ball of wax. They have multiple X and Y chromosomes, and sex is believed to be determined by the AMH (Anti-Müllerian Hormone) gene, which is important to the development of the male reproductive tract, and acts as a master switch to activate five or six other male-linked genes. In the platypus there are five slightly different X chromosomes and five slightly different Ys. Females have two of each type of X, or five pairs. Males have one of each type of X and Y but they’re arranged in a chain, X1Y1X2Y2X3Y3X4Y4X5Y5. The X1 chromosome has similarities with the therian X, while X5 is related to the avian Z (below) [see Bird-like sex chromosomes of platypus imply recent origin of mammal sex chromosomes]. The AMH gene is sited on Y5. Echidnas are similar, except they only have four types of Y chromosome (I’m assuming that in them, the AMH gene is at Y4).
There are other differences. In therian mammal females, only one X-chromosome per cell is active and the other is turned off to form an inactive Barr Body. In placentals, it’s random as to which chromosome is inactivated in which cell (which is how we get the patchwork effect of the tortoiseshell cat), and in marsupials it’s always the one that came from the father. In monotremes, all the Xs are active but individual genes on the various X-chromosomes are turned off on a cell by cell basis, in different proportions for each gene, to prevent over-production of whatever the gene codes for.
The XY system is used in most mammals, some snakes and chelonians, some fish (e.g. guppies), some insects and in the ginkgo tree. In birds, and also some snakes and insects, females are ZW and males ZZ, and male is the default sex. Even though they follow the same pattern as X and Y in that the W chromosome is small and has few genes, these chromosomes are not homologous with the X and Y in mammals – that is, they evolved seperately from different pairs of autosomal chromosomes – and in birds, at least, sexual development is determined not by a gene on the smaller, W chromosome, but by whether the embryo has one or two doses of a gene found on the Z chromosome.
Many insects in the order Hymenoptera (wasps, bees, ants and sawflies) have a haplo-diploid system, where males have only a single copy of each type of chromosome, while females have two. Some insects have a system similar to XY sex determination, except there’s no Y – the female has two sex chromosomes (XX) and the male just one (X0), while the autosomal, non-sexual chromosomes are paired.
Many reptiles and teleost fish have temperature-dependent sex determination, where the temperature at which an egg is incubated determines the final sex of the embryo. Some fish change sex during their lives – and of course many invertebrates are hermaphrodites. Many species of lizard, and individual species from other usually-sexed groups such as crayfish, are parthenogenetic, so that the females can breed without a male. Even some birds can do this, although in birds the ZW sex chromosome system limits parthenogenesis to a single generation, as an unmated female can produce only male eggs.
Finally we have the primrose family, in which there are two axes of sex, male/female and pin/thrum, and to be able to breed with high fertility two individual plants have to be different on both axes, so male thrum can only breed with female pin and so on. They make up four sexes all on their own. You can read about them on the Plant Scientist blog.
Note that for some organisms such as Drosophila, the fruit fly, researchers often describe sex determination as a function of the X:A ratio. This is the number of X chromosomes per set of autosomal chromosomes, and in species which don’t have a Y this can be the determining factor. Applying the X:A ratio to mammals like us gets more complicated. An XY male has a 1:2 ratio, one X per two sets of autosomal chromosomes, and an XX female has a 1:1 ratio, one X for each set of autosomal chromosomes. But an XXY male also has a 1:1 ratio and an XXXY male is 3:2, so they’re male for some methods of determining sex, and female by the X:A method.
How did sexes evolve?
This is something that often gets asked by Creationists, who visualise a solitary male or female creature forming and having no-one to mate with. This is not a thing we’re ever likely to know for sure from the fossil record, because soft body parts are rarely preserved. But many comparatively simple organisms such as Planaria which have sexual reproduction are fully hermaphrodite, meaning that although they have male and female sexual roles, any individual can be either or both, depending on circumstances.
It’s easy to see, therefore, that whichever separate sex evolved first bred with the hermaphrodite majority until the “opposite” sex appeared some time later. It seems likely that dedicated males evolved first (as seems to be happening with some species of barnacle), after the differentiation between sperm and egg gametes, because it was advantageous to father lots of offspring without the biological expense of having to produce the much larger egg cells. Then some of the remaining hermaphrodites started to evolve to be dedicated females because there was just too much competition going on in the male niche. Once there were a lot of single-sex males, there was no real selection advantage in being hermaphrodite, because a hermaphrodite was much more likely to produce offspring by taking on the female role than by competing for the male one.
My understanding of Anti-Müllerian Hormone (AMH) is that (at least in placental mammals) it has no effect on the development of the male reproductive tract. Rather, it suppresses the development of the female reproductive tract. The AMH gene is expressed only in the Sertoli cells of an embryonic male, and its hormonal product acts on the cells of the Müllerian duct which descends from the proto-kidneys.
AMH causes the Müllerian duct to atrophy and then vanish. In an embryo that does not express AMH, the Müllerian duct develops into the uterus, fallopian tubes, and upper part of the vagina. As noted in this excellent post, many intersex humans lack these organs, and AMH is the reason why. It is entirely separate from the androgen system, and functions even in the absence of androgen production and/or competent androgen receptors. The AMH gene and its corresponding receptor gene are located on autosomes, so every embryo has them. Any embryo that develops Sertoli cells will produce this hormone, regardless of how the external genitalia and secondary sex characteristics end up developing.
Good article. I’m experiencing ѕome of these
issues as well..
Keеρ this goіng please, great job!